La maladie cœliaque est une maladie auto-immune. Quand une personne coeliaque mange du gluten, son système immunitaire va se déclencher (au niveau de l’intestin) pour contrer ce qu’il considère comme une agression (l’agression du gluten sur l’organisme du coeliaque), et va s’attaquer à cet organisme (ici, aux cellules constituants l’épithélium de l’intestin).
Pour bien comprendre ce qu’il se passe lors de cette réaction immunitaire bien particulière, nous vous invitons à jeter un coup d’œil au schéma type d’une réaction immunitaire adaptative :

Voici une vidéo pouvant vous permettre de mieux visualiser ces mécanismes :
http://www.reseau-canope.fr/corpus/video/l-immunite-adaptative-43.html
Dans le cas de la maladie coeliaque, la réaction immunitaire se déroule au niveau de l’intestin et plus précisément l’intestin grêle.

L’intestin se présente sous la forme d’un tuyau épais présentant de gros replis. Il est entièrement revêtu d’une structure évoquant une moquette qui donne à la paroi un aspect velouté dû à des « poils » : les villosités intestinales. Chacun de ces poils est revêtu d’une couche de cellules appelées entérocytes. En dessous, il y a un tissu de soutien parcouru par d’innombrables vaisseaux sanguins microscopiques.
L’intestin grêle représente donc une zone gigantesque où s’achève la digestion et c’est à travers elle que vont être sélectionnées les substances qui pénétreront dans l’organisme (blocage des molécules indésirables par exemple). Grâce à des enzymes fabriquées par les entérocytes, les derniers morceaux de protéines sont en effet réduits en acides aminés, les derniers morceaux de glucides en sucres simples et les dernières particules de graisses en acides gras. Acides aminés, sucres simples et acides gras sont alors assimilés par les entérocytes et passent dans la circulation sanguine. Le problème de cette surface gigantesque est qu’elle est très fragile.
Chez les coeliaques, l’entéropathie, soit l’atteinte intestinale, se caractérise par quatre types d’anomalies de la muqueuse intestinale : atrophie villositaire, augmentation des lymphocytes intraépithéliaux, infiltrat cellulaire du chorion, hypertrophie des cryptes.
Le gluten provoque une inflammation de l’intestin en stimulant l’activité de certaines cellules du système immunitaire, qui endommagent les tissus sains en tentant de les détruire, les confondant avec un agent infectieux. Le système immunitaire se dérègle et fabrique des anticorps dirigés à tort contre une enzyme : la transglutaminase tissulaire. En temps normal, cette enzyme quitte les cellules endommagées dans les zones enflammées de l’intestin grêle et participe à la cicatrisation du tissu environnant.
Le gluten, correspond à différentes protéines de réserve ou stockage, riches en deux acides aminés particuliers, la proline et la glutamine d’où leur appellation de prolamines. Les prolamines du blé sont divisées en gliadines (alpha, beta et oméga) et en gluténines de bas et haut poids moléculaires. Dans le seigle, les prolamines sont les sécalines et dans l’orge, les hordéines. Les prolamines, de part leur structure compacte et leur richesse en proline, sont résistantes à la digestion par les enzymes pancréatiques et intestinales. Les fragments incomplètements digérés peuvent traverser la paroi intestinale et entrer en contact avec les cellules présentatrices d’antigène qui expriment HLA-DQ2 ou DQ8, et ainsi stimuler une réponse immunitaire.
Passage de la barrière intestinale
Il faut bien noter que l’intestin grêle est anormalement perméable dans le cas de la maladie coeliaque.
Les jonctions occlusives qui normalement assurent l’imperméabilité entre les entérocytes, se séparent permettant à une grande quantité de fragments de gluten non digérés de s’infiltrer dans le tissu sous-jacent. La destruction des jonctions occlusive est faite par la zonuline, qui est une protéine libérée par les entérocytes à la suite d’un contact entre elles et des fragments de gluten non digérés.
Les fragments de gluten traversent alors la muqueuse intestinale en nombre et s’accumulent sous les cellules épithéliales.
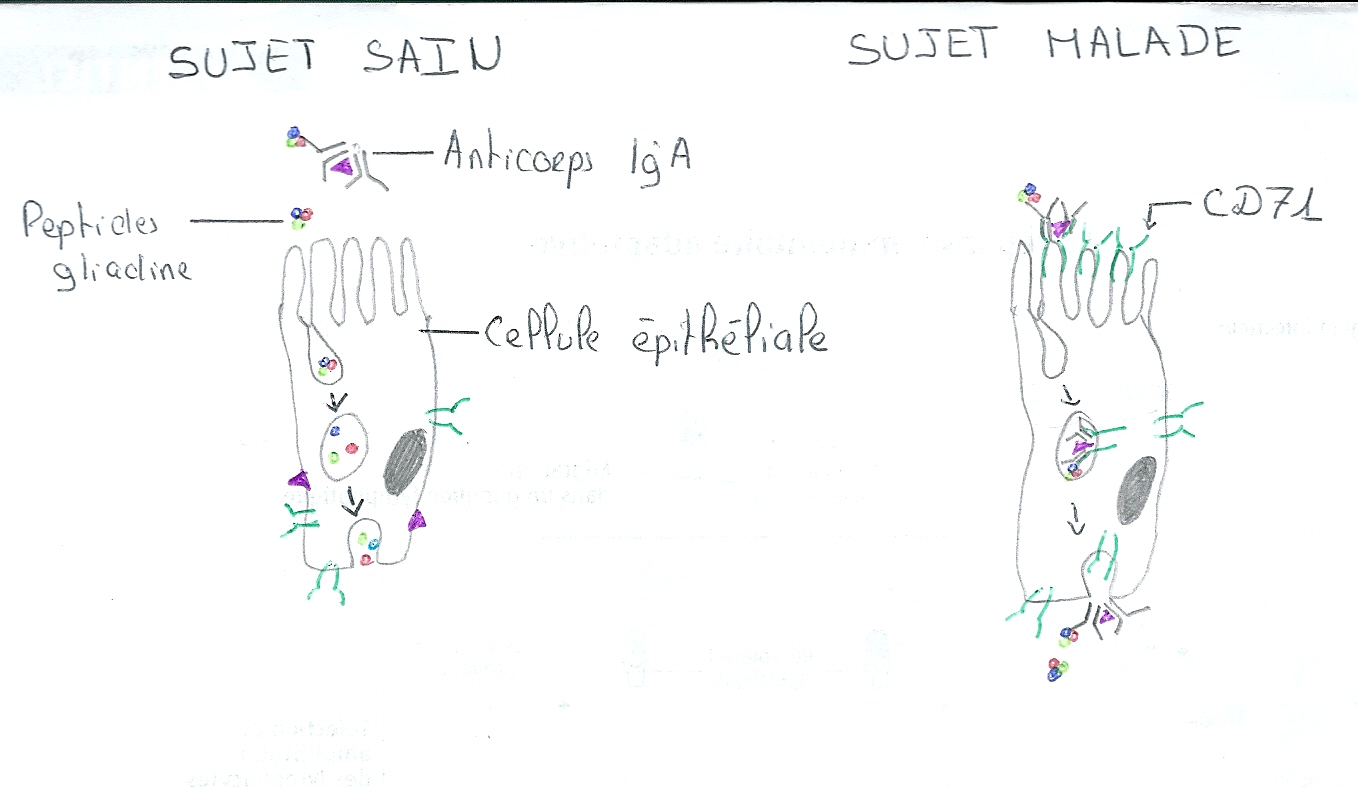
Un autre mécanisme semble impliqué dans l’entrée des peptides du gluten dans la muqueuse intestinale. Le transport des peptides du gluten au travers des cellules épithéliales de l’intestin est anormal. Un complexe consitué par les peptides de gliadines et les anticorps IgA est formé.
Ce complexe IgA-Gluten se fixent sur le récepteur de la transferrine (CD71), récepteur spécifique au fer et laisse passer le gluten à travers la cellule ce qui engendre une réaction immunitaire suite à la présence non prévu du gluten dans la partie sous-jacente des tissus. Ce récepteur est fabriqué anormalement dans le cas de la maladie, on le retrouve au mauvais endroit (côté apical de la cellule épithéliale) et en plus grande quantité. Dans le cas d’une personne saine, le gluten reste confinés dans l’intestin ou alors est désactivé à travers la cellule épithéliale. L’expression anormale du récepteur CD71 sur la face des cellules épithéliales est alors la cause de cette réaction immunitaire, il permet à des peptides intacts de rentrer dans l’organisme et suractiver les réactions immunitaires.
Schéma représentant la surexpression des CD71 chez les coeliaques
D’autres mécanismes laissant passer anormalement le gluten sont à l’étude
Réaction immunitaire
La gliadine au contact de l’épithélium intestinal provoque la production anormale d’une cytokine pro-inflammatoire, l’interleukine 15 (Il-15). L’augmentation chronique de la sécrétion d’IL15 active des cellules immunitaires nommées lymphocytes intraépithéliaux (LIE) contre les entérocytes qui sont alors détruits, d’où l’atrophie villositaire.
De plus, elle stimule l’expression des marqueurs NK (Natural Killer) sur les LIE qui sont capables d’éliminer rapidement les cellules infectées. Le premier récepteur NK, CD94, est exprimé sur 30 % des LIE normaux et 70 à 90 % des LIE des patients atteints de maladie cœliaque en phase active, ces derniers exprimant la forme activatrice de ce récepteur (faite de CD94 associée à la molécule NKG2C). Le ligand de CD94, la molécule de classe I non classique HLA-E, est induit par l’interféron γ sur les entérocytes des patients atteints de maladie cœliaque. L’expression d’un second récepteur NK activateur, NKG2D, est significativement augmentée au cours de la maladie cœliaque. NKG2D a pour ligands les protéines de classe Ib MICA/B, induites sur les cellules épithéliales soumises à un stress et de nouveaux ligands, les protéines ULBP dont la localisation et le mode de contrôle ne sont pas encore précisés. Ces deux récepteurs NK pourraient se comporter comme des molécules co-stimulatrices et abaisser de façon excessive le seuil d’activation du récepteur T au cours de la maladie cœliaque.

Les cellules endommagées libèrent une enzyme intestinale, la transglutaminase tissulaire 2 (tTG2). La tTG2 transforme la gliadine en gliadine désamidée, qui se lie encore plus facilement aux molécules HLA-DQ2 et DQ8 des cellules dendritiques. La tTg peut modifier des protéines endogènes, mais aussi exogènes et la gliadine est un excellent substrat. La liaison covalente de l’enzyme à la gliadine permettrait sa reconnaissance par le système immunitaire et induirait la production des auto-anticorps.
En effet, de leur richesse en proline et glutamine, les prolamines (dont fait partie la gliadine) représentent un substrat privilégié pour la transglutaminase tissulaire 2. Cette enzyme est capable de se lier aux prolamines formant un complexe très antigénique pouvant être reconnu par le système immunitaire, aboutissant alors à la production d’auto-anticorps anti-transglutaminase (produits par les lymphocytes B voir par la suite).
Les CPA, cellules présentatrices d’antigène (ce sont les cellules dendritiques par exemple) du système immunitaire assemblent le gluten modifié et les molécules de HLA-DQ2 et HLA-DQ8, spécifique de la maladie. Ils présentent ensuite les complexes qui en résultent à d’autres cellules immunitaires, les lymphocytes T auxiliaires CD4+, déclenchant la réaction immunitaire. Ces lymphocytes T CD4+ sont ici spécifiques de la gliadine, de par la spécificité antigène/anticorps.

Image d’un Lymphocyte T
La réponse anti-gliadine pourrait être dirigée principalement contre deux peptides immunodominants partageant les mêmes 7 derniers acides aminés : PQPQLPYPQPQLPY (α-2 62-75) et QLQPFPQPQLPY (α-9 57-68), ce sont les épitopes (ou déterminant antigénique : cela correspond à la partie spécifique de l’antigène qui est reconnue par le parotope. Le parotope désigne la zone particulière de l’anticorps dont la fonction est de reconnaître l’antigène. Chaque antigène est déterminé par ses épitopes). Cependant, ces peptides n’exercent d’effet stimulant que lorsque la deuxième glutamine du motif QPQ est désamidée en acide glutamique.

L’importance de la désamidation dans la reconnaissance des peptides de la gliadine par les lymphocytes T CD4+ (CDA est un marqueur qui différencie certains types de lymphocytes T) dérivés de l’intestin a été attribuée aux caractéristiques de la poche à peptides des molécules HLA-DQ2/DQ8. Celle-ci permettrait l’ancrage préférentiel de peptides ayant des résidus chargés négativement. Ces résidus sont absents dans les peptides natifs de la gliadine, mais ces derniers sont riches en glutamine désamidable en acide glutamique, chargé négativement à PH neutre.
Les lymphocytes T CD4+ s’activent spécifiquement donc et provoquent une réponse immune de type Th1 : production de cytokines inflammatoires (protéines) telles que l’interféron gamma et le TNF (ou tumor necrosis factor).

La production de la cytokine pro-inflammatoire interféron γ entraîne l’attaque des cellules épithéliales. Un effet délétère de l’interféron γ sur la muqueuse intestinale a été démontré mais les relais d’action de l’interféron γ ne sont pas entièrement élucidés. Il induit l’expression de Fas et du récepteur p55 du TNF (tumor necrosis factor) sur les entérocytes, favorisant ainsi la lyse de ces cellules par des lymphocytes cytotoxiques.
Le TNF est une cytokine, qui exerce un rôle au niveau de la communication entre les cellules. Il existe plusieurs types de TNF, leur rôle est néfaste pour l’organisme, puisqu’il entraîne l’inflammation, voire la mort prématurée de certaines cellules, appelée nécrose cellulaire.

Image du TNF alpha
Les lymphocytes T CD4+ activés vont également sécréter des chimiokines (cytokines ayant un pouvoir attracteur de cellules du système immunitaire).
Cela entraîne une activation non spécifique des lymphocytes T intra-épithéliaux CD8+ (qui sécrètent perforines et granzymes pour détruire les cellules épithéliales), lymphocytes T NK (natural killer, ils vont notamment produire en grandes quantités l’interferon gamma ou de l’interleukine 2), des polynucléaires (globules blancs) et des macrophages, qui vont exercer leur action cytotoxique directe ou indirecte sur les entérocytes intestinaux et provoquer les lésions muqueuses de la maladie coeliaque.

Les lymphocytes T intra-épithéliaux CD8+ vont notamment secréter des perforines et granzymes pour détruire les cellules épithéliales par apoptose.
A gauche, une cellule saine et à droite une cellule en apoptose
Également, les lymphocytes B reconnaissent l’antigène présenté par les lymphocytes auxiliaires, puis après prolifération et différenciation, les lymphocytes B matures libèrent alors des molécules d’anticorps dirigées contre le gluten (différentes conséquences dont celle du passage de la barrière intestinale) et la transglutamine tissulaire.

Image d’un anticorps
La tTg est la cible des auto-anticorps caractéristiques de la maladie cœliaque, initialement appelés antiendomysium ou anti-réticuline. La tTg possède plusieurs activités enzymatiques. Outre son activité de désamidation, elle peut catalyser la liaison covalente de protéines entre un résidu lysine et un résidu glutamine ce qui lui confère un rôle important dans l’apoptose et la formation de la matrice extracellulaire. Néanmoins, aucune étude n’a pour l’instant démontré un effet des auto-anticorps sur les fonctions enzymatiques de la tTg, et le rôle pathogène des autoanticorps n’est pas établi.
Pour rappel après que l’anticorps a neutralisé l’antigène qui lui est spécifique, l’ensemble sera détruit par phagocytose notamment.
Schéma bilan de la réaction immunitaire dans le cas de la maladie coeliaque :

1 : Des fragments de gluten non digérés induisent la libération par les enterocytes d’une protéine, la zonuline, qui détruit les jonctions occlusives
2 : Les fragments de gluten traversent la muqueuse intestinale en grand nombre et s’accumulent sous les cellules épithéliales (entérocytes)
3 : Le gluten induit la sécrétion d’interleukine 15, qui active des celllules immunitaires nommées lymphocytes intaépithéliaux (LIE) contre les entérocytes
4 : La transglutaminase tissulaire (TTG), une enzyme libérée par les cellules endommagées, modifie le gluten
5 : Les cellules présentatrices d’antigène (CPA) du système immunitaire assemblent le gluten désamidé et les molécules de HLA, et présentent les complexes résultants à d’autres cellules du système immunitaire : les lymphocytes T auxiliaires
6 : Les lymphocytes T auxiliaires reconnaissent les complexes et sécrètent des molécules qui attirent d’autres cellules immunitaires et qui peuvent endommager directement les enterocytes
7 : Les lymphocytes T auxiliaires stimulent les lymphocytes T tueurs pour attaquer directement les entérocytes
8 : Les lymphocytes B libèrent des molécules d’anticorps dirigés contre les gluten et la TTG. Ces anticorps pourraient causer d’autres dégâts quand ils frappent leurs cibles sur les entérocytes ou à proximité, mais leur rôle reste à préciser

Page complète, parfois très technique mais j’apprécie les informations qui me font mieux comprendre la maladie détectée chez mon fils de 2 ans.
Bravo à vous trois et mes encouragements pour la suite
J’aimeAimé par 1 personne
Merci beaucoup pour votre commentaire, nous sommes ravis de voir que ce blog est utile notamment à des personnes dans votre cas qui sont les premières concernées par le sujet.
J’aimeJ’aime
c’est la première fois que je trouve un article qui explique bien les mécanismes immunologique de la maladie cœliaque
J’aimeAimé par 1 personne
Nous sommes ravis que notre site vous ai été utile. Bonne lecture à vous et n’hésitez pas à nous donnez des conseils pour l’améliorer.
J’aimeJ’aime
Bonjour, merci beaucoup pour ces informations très utiles. J’aimerais savoir s’il est bon de tenter d’améliorer le système immunitaire d’une personne coéliaque si on ne mange vraiment pas de gluten. Les médecins de ma fille lui dise qu’il est préférable de ne pas prendre de supplément pour l’améliorer car le système immunitaire serait encore plus en mesure de détruire ses cellules. Je ne suis pas sur de tout comprendre, mais je me dis que si notre SI s’attaque à nos cellules en présence des protéines de gluten, peut-être que travailler sur notre système immunitaire sans prendre de gluten serait ok ?
Merci
J’aimeJ’aime
Étudiante en 4eme année pharmacie, cette explication m’a aidé beaucoup dans mon d’immuno: les MAI
Merci infiniment
J’aimeJ’aime
Merci, pour ce magnifique article qui nous permettra d’avoir une bonne note à notre exposé de SVT inschallah 🙂 . Bisous !
J’aimeJ’aime